4 - Relationship to other sensory systems
According to textbooks on comparative anatomy , the
origin of auditory systems in higher vertebrate species
can be traced to the lateral line organs of fish [1, 2].
The lateral line system evolved from neural
mechanisms for orientation. Later elaborations
provided functions for the vestibular system of animals
adapted to living on land. The inferior colliculus of
higher vertebrate species has analogous functions to
the vestibular organ (torus semicircularis) in the brain
of lower vertebrates [3].
In mammals, the vestibular sense organs reside in the
inner ear and there are neural connections from
auditory nuclei to subcortical components of the
vestibular system. Thus auditory function remains
closely linked to vestibular function, and signs of
vestibular dysfunction have been reported in autistic
children [4].
Orientation and response to vibrations transmitted
through water may be the earliest precursors of
auditory function in primitive species of fishes.
However, the inferior colliculi evolved as a posterior
extension of the optic lobe in amphibian species. The
anterior part of the optic lobe evolved into the superior
colliculi in higher vertebrates. The superior and
inferior pairs together are known as the corpora
quadrigeminae, and are situated in the tectum (roof) of
the midbrain above the third ventricle of the brainstem,
just forward of the cerebellum. These structures are
depicted in figures 9 and 11.
The superior colliculi detect change in the peripheral
visual fields. Detection of motion in the visual field has
long been recognized as important for survival. Lettvin
et al. (1959) described the firing of neurons in the
optic lobe of frogs in response to movement in the
visual field [5]. The inferior colliculi added detection of
acoustic change in larger and more distant extents
within the surroundings of later developing species.
Denny-Brown (1962) proposed that the optic tectum is
the most essential structure in the brain for unitary
behavior [6]. With extensive connections to other
sensory and motor systems of the brain, the inferior
colliculi should be considered, along with the superior
pair, as possible locus of the conscious state.
But why should the inferior colliculus have a higher
metabolic rate than the superior colliculus? Perhaps
this is because, as an accessory to the primary visual
alerting system, the inferior colliculus provides not just
a new set of functions for hearing but also
mechanisms that make use of auditory and visual
signals simultaneously and in a way that the two
systems can interact. Hearing is an accessory to
vision in the species familiar to most of us. It would be
interesting to find a species in which hearing evolved
first with the visual system as the later-developing
elaboration. The auditory system may have more work
to do thus require more energy than if it evolved
without any relationship to the visual system. We turn
our heads and look for things that make noises but
rarely try to hear something that happens to pass by
our field of vision.
According to textbooks on comparative anatomy , the
origin of auditory systems in higher vertebrate species
can be traced to the lateral line organs of fish [1, 2].
The lateral line system evolved from neural
mechanisms for orientation. Later elaborations
provided functions for the vestibular system of animals
adapted to living on land. The inferior colliculus of
higher vertebrate species has analogous functions to
the vestibular organ (torus semicircularis) in the brain
of lower vertebrates [3].
In mammals, the vestibular sense organs reside in the
inner ear and there are neural connections from
auditory nuclei to subcortical components of the
vestibular system. Thus auditory function remains
closely linked to vestibular function, and signs of
vestibular dysfunction have been reported in autistic
children [4].
Orientation and response to vibrations transmitted
through water may be the earliest precursors of
auditory function in primitive species of fishes.
However, the inferior colliculi evolved as a posterior
extension of the optic lobe in amphibian species. The
anterior part of the optic lobe evolved into the superior
colliculi in higher vertebrates. The superior and
inferior pairs together are known as the corpora
quadrigeminae, and are situated in the tectum (roof) of
the midbrain above the third ventricle of the brainstem,
just forward of the cerebellum. These structures are
depicted in figures 9 and 11.
The superior colliculi detect change in the peripheral
visual fields. Detection of motion in the visual field has
long been recognized as important for survival. Lettvin
et al. (1959) described the firing of neurons in the
optic lobe of frogs in response to movement in the
visual field [5]. The inferior colliculi added detection of
acoustic change in larger and more distant extents
within the surroundings of later developing species.
Denny-Brown (1962) proposed that the optic tectum is
the most essential structure in the brain for unitary
behavior [6]. With extensive connections to other
sensory and motor systems of the brain, the inferior
colliculi should be considered, along with the superior
pair, as possible locus of the conscious state.
But why should the inferior colliculus have a higher
metabolic rate than the superior colliculus? Perhaps
this is because, as an accessory to the primary visual
alerting system, the inferior colliculus provides not just
a new set of functions for hearing but also
mechanisms that make use of auditory and visual
signals simultaneously and in a way that the two
systems can interact. Hearing is an accessory to
vision in the species familiar to most of us. It would be
interesting to find a species in which hearing evolved
first with the visual system as the later-developing
elaboration. The auditory system may have more work
to do thus require more energy than if it evolved
without any relationship to the visual system. We turn
our heads and look for things that make noises but
rarely try to hear something that happens to pass by
our field of vision.
- Ariens Kappers CU et al.
(1936) The Comparative
Anatomy of the Nervous
System of Vertebrates
including Man. - Sarnat HB, Netsky, MG (1974)
Evolution of the Nervous
System. - Heiligenberg W, Rose G
(1985)Phase and amplitude
computations in the midbrain
of an electric fish: intracellular
studies of neurons
participating in the jamming
avoidance response of
Eigenmannia. - Ornitz EM (1983) The
functional neuroanatomy of
infantile autism. - Lettvin JY et al. (1959) What
the frog's eye tells the frog's
brain. - Denny-Brown D (1962) The
midbrain and motor
integration.
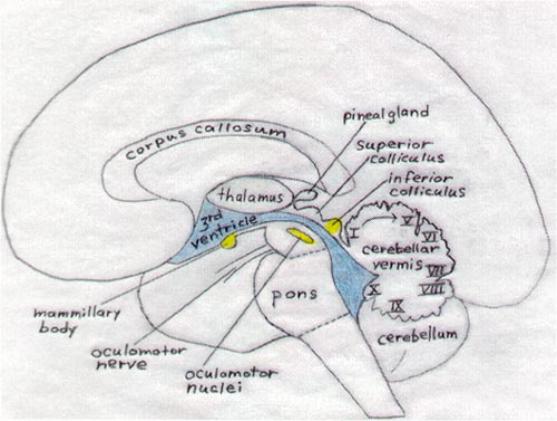
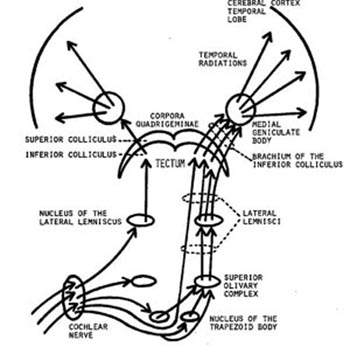
Figure 11 -
Diagram of the
auditory system.
Diagram of the
auditory system.
- Ariens Kappers CU, Huber GC, Crosby EC (1936) The Comparative Anatomy of the
Nervous System of Vertebrates including Man. New York: MacMillan. - Sarnat HB & Netsky, MG (1974) Evolution of the Nervous System New York: Oxford
University Press. - Heiligenberg W, Rose G (1985)Phase and amplitude computations in the midbrain of
an electric fish: intracellular studies of neurons participating in the jamming
avoidance response of Eigenmannia. Journal of Neuroscience 5:515-531 - Ornitz EM (1983) The functional neuroanatomy of infantile autism. International
Journal of Neuroscience 19:85-124. - Lettvin JY, Maturana H, McCulloch WS, Pitts WH (1959) What the frog's eye tells the
frog's brain. Proceedings of the Institute of Radio Engineers 47:1940-1951. - Denny-Brown D (1962) The midbrain and motor integration. Proceedings of the Royal
Society of Medicine 55:527-538.